可视化理解光合作用
光合作用(Photosynthesis)是地球上生命的一项基本且至关重要的过程,它主要由植物、某些细菌和藻类执行。利用光能将水、二氧化碳或硫化氢等无机物转变成可以储存化学能的有机物(比如碳水化合物)的生物过程,是地球生物圈主要的初级生产模式。简单理解,光合作用即采集阳光制造糖类!♻️
光合作用的原理
光合作用文字方程式:二氧化碳 + 水 + 光能 ⟶ 葡萄糖 + 氧气 + 水
这个过程的关键参与者是细胞内部的叶绿体。叶绿体在阳光的作用下,把经由气孔进入叶片内部的二氧化碳和由根部吸收的水转变成为葡萄糖,同时释放出氧气:
$12 H_2O + 6 CO_2 \rightarrow C_6H_{12}O_6(葡萄糖)+ 6 O_2 + 6 H_2O$
植物的光合作用可分为光反应和碳反应两个步骤如下:
$12 H_2O + 阳光 \rightarrow 24 H^+ + 6 O_2 + 12e^- \ [光反应]$
$24 H^+(来自光反应)+ 6 CO_2 \rightarrow C_6H_{12}O_6(葡萄糖) + 6 H_2O \ [碳反应]$
叶绿体

叶绿体是三种类型的质体(plastid)(其他两个质体类型是白色体和有色体,含有少量叶绿素并且不能进行光合作用)之一,其特点是具有高浓度的叶绿素。叶绿体是高度动态的,它们在植物细胞内四处移动,并且偶尔分裂。它们的行为受到环境因素(如光的颜色和强度)的强烈影响。叶绿体和线粒体类似,拥有自身的遗传物质DNA,但因其基因组大小有限,所以是一种半自主细胞器。其DNA被认为是从已被古代真核生物的细胞吞没的有光合作用的蓝菌门祖先继承下来。叶绿体不能由植物细胞产生,且必须在植物细胞分裂期间由每个子细胞继承叶绿体。
💡 注意
能进行光合作用的生物,不一定有叶绿体。例如,蓝细菌是原核生物,没有叶绿体,但因其含有叶绿素和藻蓝素,也可进行光合作用。
光反应
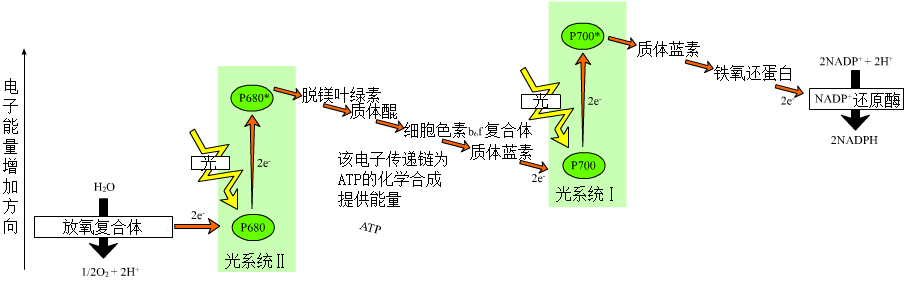
光反应是反应中心色素所吸收的光能与原初电子受体和次级电子受体之间进行的氧化还原反应,从而实现光能转化为电能,并转变为化学能的过程。这个过程主要是光合磷酸化【以阳光作为能量源,将ADP磷酸化为ATP的过程】,在光合磷酸化中,光能用于产生高能电子供体和低能电子受体。然后,电子通过电子传输链【电子从高能分子(供体)到低能分子(受体)的转移可以在空间上被分为一系列中间的氧化还原反应】自发地从供体向受体移动产生能量【一系列氧化还原反应被用来产生跨膜的电化学势梯度或质子动力(pmf),这种跨膜电化学势梯度的形式产生了能量】。最后,用来产生ATP和NADPH。
- 场所:类囊体膜
- 影响因素:光强度,水分供给,氧含量
- 详细过程:
- 叶绿体膜上有两套光合作用系统(光系统):光合作用系统I和光合作用系统II【光合作用系统I比光合作用系统II要原始,但电子传递先在光合作用系统II开始,一、二的命名则是按其发现顺序】。
- 在光照条件下,两系统分别吸收
700nm和680nm波长的光子作为能量,将从水分子在光解过程中得到的电子不断传递,该过程还有细胞色素b6f的参与。 - 电子最后被传递给辅酶NADP,通过铁氧还蛋白-NADP还原酶将NADP还原为NADPH。
- 而水光解所得的氢离子则顺浓度差通过类囊体膜上的蛋白质复合体而从类囊体内向外移动到基质,势能降低。
- 其间的势能用于合成ATP【由一种称为ATP合酶的酶合成的,ATP合酶由跨膜电化学势梯度(通常以质子梯度的形式存在)驱动】,以供碳反应所用。
- 此时势能已降低的氢离子则被氢载体NADP带走,一分子NADP可携带两个氢离子。这个NADPH与氢离子则在碳反应中充当还原剂的作用。
- 反应产物:
- 光解水,产生氧气。
- 将光能转变成化学能,产生三磷酸腺苷 (ATP),为碳反应提供能量。
- 利用水光解的产物氢离子,合成NADPH及氢离子,为碳反应提供还原剂。
细说下这种光合磷酸化有两种反应途径:非循环光合磷酸化【非循环电子传递链】和循环光合磷酸化【循环电子传递链】。
在类囊体膜上的非循环电子传递链过程:光系统II ⟶ 初级接受者(Primary acceptor) ⟶ 质体醌(Pq) ⟶ 细胞色素复合体(Cytochrome Complex) ⟶ 质粒蓝素(含铜蛋白质,Pc) ⟶ 光系统I ⟶ 初级接受者 ⟶ 铁氧化还原蛋白(Fd) ⟶ NADP + 还原酶(NADP + reductase)
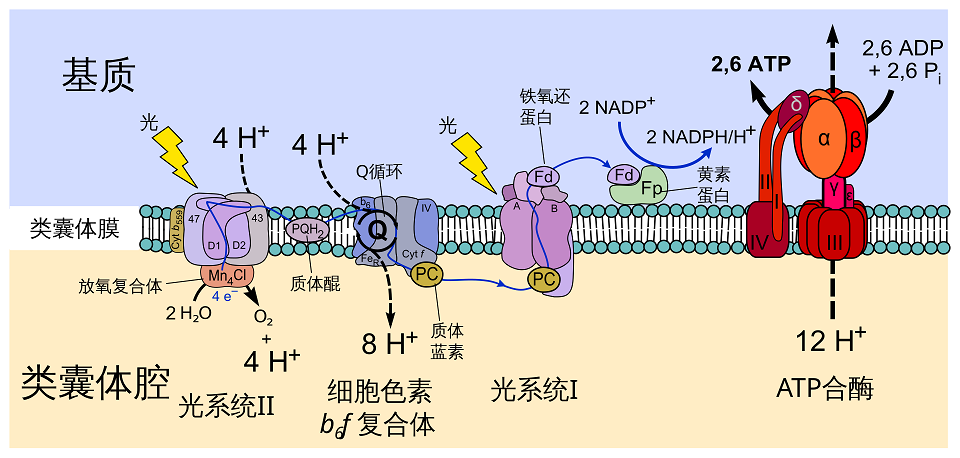
如上图举例说明电子传递:从光系统II出发后,会光解水,释放氧气,ATP与NADPH。
- 一个水分子被分解成${4 H^+ + O_2 + 4 e^-}$,这个过程被称为光解(或光分裂)。其中的4个电子保留在光系统II中,而${4 H^+}$和${O_2}$则留作之后使用。
- 光系统反应核心中心周围的光合色素吸收一个光子。光激发这些色素中的电子,引起连锁反应,最终将能量转移到光系统II的核心,并激发主要电子受体脱镁叶绿素的4个电子。电子的不足由另一个水分子中的电子来补充。
- 电子从脱镁叶绿素传输到质体醌,它从脱镁叶绿素接受${4 e^-}$,从基质接受${4 H^+}$离子并合成${PQH_2}$。
- 它后来被分解成${PQ}$,将4个电子释放到细胞色素
b6f复合,8个${H^+}$离子释放到类囊体腔中。 - 电子通过
Cyt b6和Cyt f。然后,它们被传递到质体蓝蛋白中,这个过程为氢离子${H^+}$提供能量,以将其泵入类囊体空间。 - 它产生了一个梯度,使${H^+}$离子流回到叶绿体的基质中【仍然被激发的电子被转移到一个光系统I化合物中】。
- 第二个来自太阳的光子将它们的能级提高到一个更高的水平。高激发电子被转移到一个受体分子上,这个受体被称为铁氧还蛋白$-NADP^+$还原酶,该酶利用它们来催化反应:${2NADP^+ + 4H^+ + 4e^- \rightarrow 2NADPH + 2H^+}$,这个反应消耗了水分解产生的${H^+}$离子。
- 最终,这个反应消耗了光子和水,并生成了$O_2$、$2ATP$、$2NADPH$与${2H^+}$。
那么,再看循环电子传递链的过程:光系统I ⟶ 初级接受者(Primary acceptor) ⟶ 铁氧化还原蛋白(Fd) ⟶ 细胞色素复合体(Cytochrome Complex) ⟶ 质粒蓝素(含铜蛋白的循环电子传递链不会产生氧气,因为电子来源并非是水的光解),最后会生产出三磷酸腺苷(ATP) ⟶ 光系统I
这种形式的光合磷酸化作用发生在叶绿体的基质层或品格通道上。在循环光合磷酸化中,从光系统I的P700【700nm波长光】释放的高能电子顺着一条环状路径流动。在循环电子流中,电子从称为光系统I的色素复合物开始,从主要受体传递至铁氧还蛋白,接着传递至质体醌 ,然后传递到细胞色素b6f(它类似于线粒体中发现的化合物),最后通过质体蓝素返回光系统I。该运输链产生质子动力,以将$H^+$离子泵到膜外。这产生了一个浓度梯度,它在化学渗透过程中用来驱动ATP合酶。这条反应路径被称为循环光合磷酸化,它既不产生$O_{2}$也不产生NADPH。与非循环光合磷酸化不同,电子没有被$NADP^+$吸收,而是被送回细胞色素b6f复合体。
碳反应
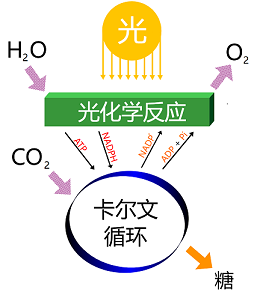
固碳作用实质上是一系列酶促反应的结果。生物界有几种固碳方法,其中以卡尔文循环为主,但并非所有进行光合作用的细胞都使用卡尔文循环进行碳固定【例如绿硫细菌会使用还原性三羧酸循环,绿曲挠菌(Chloroflexus)会使用3-羟基丙酸途径(3-Hydroxy-Propionate pathway),还有一些生物会使用核酮糖-单磷酸途径(Ribolose-Monophosphate Pathway)和丝氨酸途径(Serine Pathway)进行碳固定】。
- 场所:叶绿体基质
- 影响因素:温度,二氧化碳浓度
- 过程:不同的植物,固碳作用的过程不一样,而且叶片的解剖结构也不相同。这是植物对环境的适应的结果。固碳作用可分为C3,C4和CAM(景天酸代谢)三种类型。三种类型由二氧化碳固定过程的不同而划分。
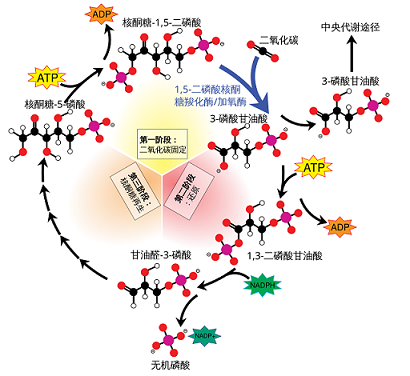
卡尔文循环详细可分为三个阶段:
- 二氧化碳固定【羧化】:大部分植物会将吸收到的一分子二氧化碳,通过核酮糖-1,5-二磷酸羧化酶/加氧酶的作用,整合到一个五碳糖分子1,5-二磷酸核酮糖(RuBP)的第二位碳原子上。此过程称为二氧化碳的固定。这一步反应的意义是,把原本并不活泼的二氧化碳分子激活,使之随后能被还原。
- 还原:但这种六碳化合物极不稳定,会分解为两分子的三碳化合物3-磷酸甘油酸。后者被在光反应中生成的NADPH与氢离子还原,此过程需要消耗ATP。产物是3-磷酸丙糖。
- 二磷酸核酮糖的再生。后来经过一系列复杂的生化反应,一个碳原子将会被用于合成葡萄糖而离开循环。剩下的五个碳原子经一系列变化,最后再生成一分子1,5-二磷酸核酮糖,循环重新开始。
💡 注意
卡尔文循环每进行六次,生成一分子葡萄糖。
光合作用的过程
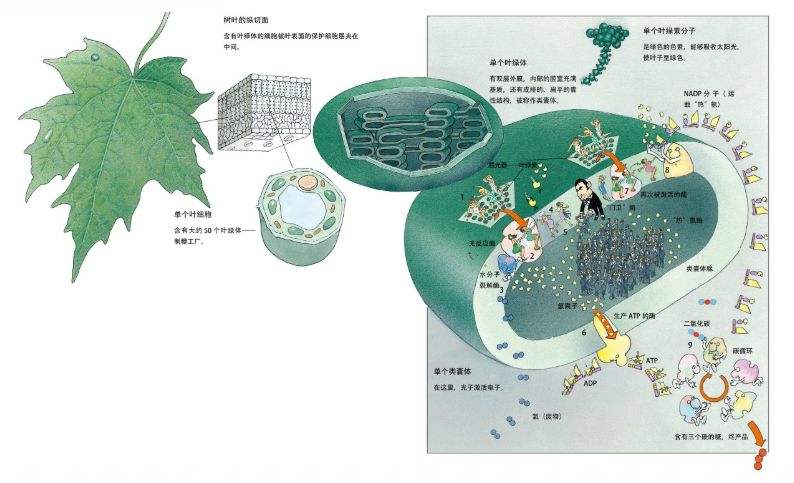
先简略介绍绿色植物生产糖的过程:
- 1、富含能量的光子打在叶片里的叶绿素上,叶绿素分子里的电子即被激发跳到能量更高的轨道上
- 2、这些精力充沛的电子,连续跳过一串叶绿素分子(跳着摇摆舞的女人),最后落在一些更小的载体分子上(其他的女舞者)
- 3、叶绿素分子丢失的电子由水分子分解释放的电子来补充,然后叶绿素会再次投入战斗之中
- 4、带负电的载体分子易于吸附氢离子(男舞者),并将氢离子转交给一种蛋白质(看门的壮汉)
- 5、这种蛋白质可以把氢离子扔进叶绿体里的类囊体
- 6、类囊体内氢离子越积越多,氢离子们便急着以一种酶作为通道(旋转门),夺门而出,这个集体外流的过程能促使酶合成大量ATP
- 7、电子再次被光能激活
- 8、在一种叫作烟碱胺腺嘌呤二核苷酸(NADP)的特殊分子上与氢离子结合,形成活性极高的“热氢”
- 9、一组酶利用ATP的能量,将周围环境中二氧化碳与“热氢”结合在一起,终于生产成糖
💡 注意
据估计,一棵成年的健康枫树的树叶总面积将近50平方米,总重量约227千克。这意味着这棵枫树的叶绿体总面积为360平方千米,在晴朗的一天中,它可以制造2吨糖!😲
1、电子被光照激发
光系统由多种色素,如叶绿素a(Chlorophyll a)、叶绿素b(Chlorophyll b)、类胡萝卜素(Carotenoids)等组成。多种色素既拓宽了光合作用的作用光谱,其他的色素也能吸收过度的强光而产生所谓的光保护作用(Photoprotection)。
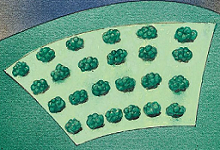
叶绿素分子成簇地排列着【这样排列提高光能捕获和转化效率,同时提供能量转移和保护机制,确保光合作用高效、安全地进行】,充当阳光感受器的角色【主要吸收可见光谱中的红色和蓝色波段(不吸收绿光,因此呈现绿色)】。
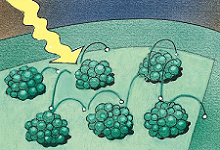
当阳光照射在叶绿素分子上时,它们的电子被激发到能量更高的轨道上,四处跳跃着,直到被一种酶捕获!
2、逃逸电子被转移
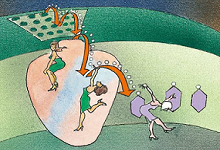
一种特殊的叶绿素——酶复合体把电子转移到类囊体膜上的载体分子中【在光系统里,当光子击中系统里的色素分子时,使电子在分子之间移转,直到到达反应中心为止】。反应中心有两种,分别位于光系统I与光系统II。光系统I的吸收光谱于700nm达到峰值,系统II则以680nm为峰值。
3、水分子被裂解

与此同时,一种裂解水分子的酶把一个水分子分解成两个电子,两个氢离子和一个氧离子【水光解作用(water photolysis)】。这里提下,氧气作为废气被排除(通过类囊体膜的孔道释放到细胞外),其他粒子被用到下面的过程中。
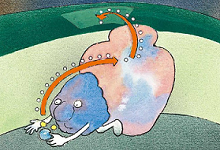
叶绿体丢失的电子将由来自水分子的电子补充【反应中心由叶绿素a及特定蛋白质所组成(这边的叶绿素a是因为位置而非结构特殊),蛋白质的种类决定了反应中心吸收光线的波长。吸收相应波长的波长的光线后,叶绿素a激发出一个电子,而旁边的酶使水光解成氢离子、电子和氧原子。此时多余的电子去补叶绿素a分子上的缺少】。
💡 注意
自然光合作用中太阳能到化学能的转换效率太低,理论最高能量转换效率约为4.6~6%,实际效率仅有1.9~2.5%
4、氢离子和电子相互吸附

因为异性相吸的原理,载体上的电子能够吸引类囊体外的氢离子。回忆一下,我们讲过一个电子加上一个氢离子就能形成一个氢原子。
5、门卫酶开始作业
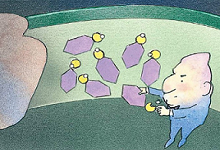
当载体到达类囊体的内膜时,“门卫”酶抓走了氢离子。

氢离子被扔进囊内,剩下的电子另有用途。
6、开始合成ATP

氢离子离开类囊体的唯一通道是膜上的一种能够生产ATP的酶【在非循环电子传递链中,细胞色素复合体会将氢离子打到类囊体(Thylakoid)里面。高浓度的氢离子会顺浓度差向类囊体外扩散。由于类囊体膜由双层磷脂分子组成,所以氢离子必须靠通道蛋白进行扩散,这个通道蛋白就是ATP合酶】。

这种酶能够提供能量,给已经消耗的ATP残体重新加上磷。
7、叶绿素复原
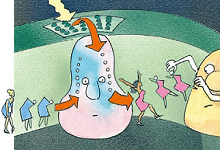
原先被剥夺了氢离子的电子被用来补充第二种叶绿素分子受光照激活而损失的电子。重获电子的叶绿素又要投入下一轮战斗了。
8、继续转移到NADP上
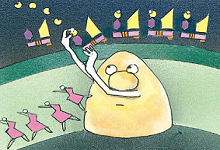
最后,一种“热”氢酶把被激活的电子和一个氢离子一起转移到最后一个载体——NADP上。
9、碳循环开始了
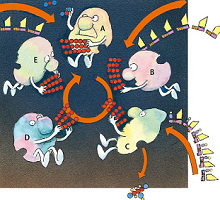
现在战场转移到了基质——在叶绿体以内,类囊体以外的地方。碳循环开始了,这个过程中有5种酶协作,利用ATP和叶绿体刚刚制作好的“热”氢生成糖的前体分子。
- 1、酶A把3个二氧化碳分别加到3个五碳糖分子上(这里没有显示氧)。生成的3个六碳糖又分解成6个三碳糖。
- 2、酶B用ATP激活这些三碳糖片段,
- 3、酶C给这6个糖分子片段加上氢,但是只把其中的一个片段踢出流水作业线。
- 4、酶D把剩下的5个三碳糖重新进行排列组合成3个五碳糖酶E用ATP把这3个糖分子激活。循环又开始了。
三种固碳方式
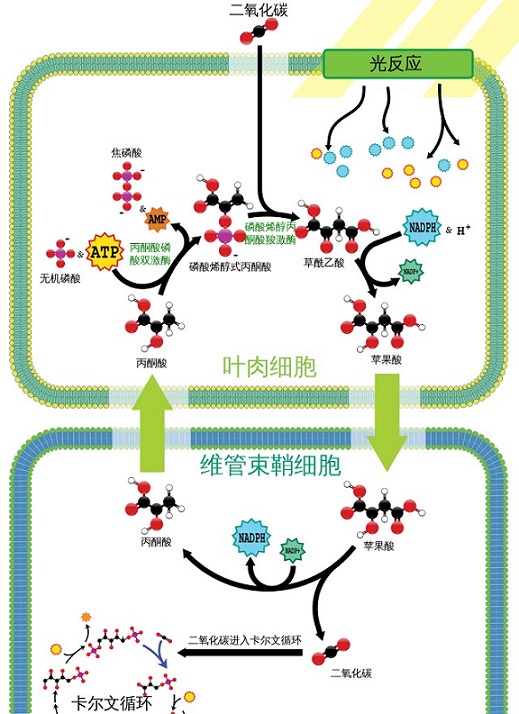
植物的碳固定方式,即植物通过光合作用将大气中的二氧化碳转化为有机物的途径,主要有以下三种:
- C3途径(卡尔文循环):这是光合作用中最普遍的一种碳固定方式,大多数植物,包括小麦、水稻、大豆等都采用这一途径。C3植物在光合作用过程中,直接由核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)催化$CO_2$与核酮糖-1,5-二磷酸(RuBP)反应,生成3-磷酸甘油酸(PGA),随后通过一系列酶促反应,转化为葡萄糖和其他有机物。C3途径的效率受光、水分和二氧化碳浓度的影响,特别是在高温和低湿条件下,由于气孔关闭减少水分蒸发,限制了$CO_2$的进入,C3植物的光合作用效率会降低。
- C4途径:C4植物,如玉米、甘蔗、熊猫竹等,通过C4途径进行碳固定,这是一种高效率且具有CO2浓缩机制的途径。在C4植物中,$CO_2$首先在叶肉细胞的PEP羧化酶作用下,与磷酸烯醇式丙酮酸(PEP)结合生成草酰乙酸,然后转化为苹果酸或天冬氨酸,通过维管系统运输到维管束鞘细胞,在这里,苹果酸或天冬氨酸释放出$CO_2$,再进入卡尔文循环。C4途径的优点是,它避免了在高温和低湿条件下的光呼吸损失,提高了光合作用效率,适合于热带和亚热带的环境。过程见上图。
- CAM途径(景天酸代谢):CAM途径是适应干旱和炎热环境的一种特别碳固定途径,主要见于仙人掌、多肉植物等。CAM植物在夜间气孔开放,吸收$CO_2$并将其固定为苹果酸或天冬氨酸,存贮于液泡中;白天气孔关闭,减少水分蒸发,释放和利用储存的$CO_2$进行光合作用。这种方式最大限度地减少了水分的流失,同时保证了光合作用的进行,是植物适应极端环境的一种策略。
这三种碳固定途径在光合作用机理上有所差异,体现了植物在不同环境条件下进化的特异适应性。
为什么是碳
碳在生命世界扮演着如此重要的角色,以至于我们可以说生命是以碳为基础的(人体内就有24%的元素含量是碳,我们常戏称为碳基生物)。这种至高无上的地位,来自碳独一无二的,能与其他原子建立起四个相互独立的化学键的本领(也就是说,碳可以贡献四个电子与其他原子共享)。氧能和其他原子形成两个键,而氢只能形成一个(另一种构建生命体的重要元素——氮占了体内原子总数的1%,它可以形成三个化学键)。我们用水管来打个比方,你就能明白碳原子那套强大的形成化学键的本领是多么无价。现在,有一款两端各有一个接口的短管子,彼此连接可以形成一根长长的水管,但仅此而已。如果短管子有三或四个接口,那它们连接成的长水管上,就有许多垂直接口可用来连接其他的物件。同样的道理,在长链大分子中,碳不仅能延长分子主干的长度,还能使主干生出侧链吸附更多基团。主干是长链分子的支撑结构,侧链基团则赋予该分子独特的化学性质和有价值的生物信息。

碳分子可以接龙:一截有四个接口的管子可以连接形成主干,而且主干上还会有分支可以连接更多其他的物件,这样一来每一段主干管道都各具特色。
顺便说下光呼吸
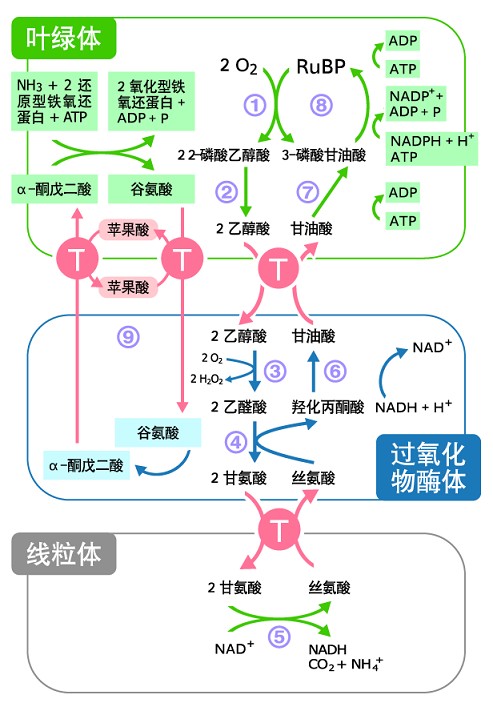
光呼吸涉及三个细胞器的相互协作:叶绿体、过氧化物酶体和线粒体。整个过程可被看作由二磷酸核酮糖被加氧分解为2—磷酸乙醇酸和3—磷酸甘油酸开始,经过一系列的反应将两碳化合物磷酸乙醇酸生成3—磷酸甘油酸,后者进入卡尔文循环,可再次生成为二磷酸核酮糖。而叶绿体内进行的是光呼吸开始和收尾的反应,过氧化物酶体内进行的是有毒物质的转换,而线粒体则将两分子甘氨酸合成为一分子丝氨酸,并释放一分子二氧化碳和氨。在光呼吸过程中产生的氨,细胞能通过谷氨酰胺—谷氨酸循环快速固定再次利用高效回收,这个过程消耗一分子ATP和NADPH。在陆生C3植物中,在光呼吸过程中产生的氨量比植物根部能吸收到的还要多,成为植物自身氮代谢的一个重要环节。而且相比起根部通过吸收硝酸根或直接从根瘤中得到氨的固定途径,光呼吸的氨固定效率要高出5到10倍。
回看研究历史
虽然一些在光合作用中的步骤仍不能被完全理解,但是整体的光合方程序自19世纪以来是已知的。
- 古希腊哲学家亚里士多德认为,植物生长所需的营养全来自土中。
- 1642年比利时人范·海尔蒙特做了柳树实验,推论植物的重量主要不是来自土壤而是来自水。但他没有发现空气中的物质也参与了有机物的形成。
- 1771年,英国的普里斯特利发现植物能够更新由于蜡烛燃烧或动物呼吸而变得污浊了的空气;但他并没有发现光的重要性。普里斯特利还发现置于密封玻璃罩内的老鼠极易窒息,但是如果加入一片新鲜薄荷叶,老鼠就可以苏醒。
- 1773年,荷兰的英格豪斯证明只有植物的绿色部分在光下才能起使空气变“好”的作用。
- 1774年,英国的普里斯特利发现绿色的植物会制造、释放出氧气。
- 1782年,瑞士的让·塞纳比耶发现,即使植物有受到阳光照射,照样会释放出氧气。
- 1804年,瑞士的索绪尔通过定量研究进一步证实二氧化碳和水是植物生长的原料。
- 1845年,德国的迈尔发现植物把太阳能转化成了化学能。[4][需要较佳来源]
- 1864年,德国的萨克斯发现光合作用会产生淀粉。
- 1880年,德国的恩格尔曼发现叶绿体是进行光合作用的场所。
- 1883年,德国的恩格尔曼运用三棱镜将太阳光折射出各色光,照射丝状海绵。一段时间后放入好氧细菌,发现植物在红光与蓝光区释放较多氧气
- 1897年,美国的Charles Reid Barnes首次在教科书中称它为光合作用。
- 1930年,C·B·凡尼尔借由对紫硫菌的研究推论植物光合作用产生的氧来自水而非二氧化碳,革新了当时的观念。
- 1941年,美国的鲁宾和卡门利用同位素标记法进行探究。证明光合作用释放氧气来自于水。
- 1945-1957年,梅尔文·卡尔文利用碳14当追踪标的,找出植物将二氧化碳转化成碳水化合物的途径。
- 1961年,彼得·米切尔发表化学渗透理论解释光反应中ATP的生成。
- 诺贝尔奖得主、科学家鲁道夫·马库斯发现电子转移链的作用和意义。
- 奥托·海因里希·瓦尔堡和迪安·伯克发现I-量子光合作用的反应,分裂二氧化碳,由呼吸作用激活。
- 2018年6月,美国《科学》杂志刊登的一项新研究说,蓝藻可利用近红外光进行光合作用,其机制与之前了解的光合作用不同。这一发现有望为寻找外星生命和改良作物带来新思路。新研究发现,上述蓝藻在有可见光的情况下,会正常利用”叶绿素a”进行光合作用,但如果处在阴暗环境中,缺少可见光,就会转为利用”叶绿素f”,使用近红光进行光合作用。
以热力学角度看
❔ 如何理解
当且仅当热力学上有利的反应同时发生时,生物大分子才能催化热力学上不利的反应。这条原理是所有已知生命形式的基础!
这条原理描绘的是生物催化剂(主要是酶)在生命活动中扮演的关键角色,以及热力学在生物化学反应中的作用。
- 热力学上的有利反应和不利反应:根据热力学第二定律,系统趋向于熵增,即自发过程倾向于向着熵增加的方向进行。在生物化学中,一个反应如果自由能变化(ΔG)为负,则是热力学上自发的,即热力学上有利的;反之,如果ΔG为正,则是非自发的,即热力学上不利的。
- 生物大分子的催化作用:生物体内的许多化学反应都是非自发的,即热力学上不利的。但是,这些反应对生物过程至关重要。生物大分子,特别是酶,具有独特的能力来降低这些反应的活化能,使得这些反应在生物体内能够发生。酶通过提供一个反应途径,使得热力学上不利的反应能够通过热力学上有利的中间步骤进行,从而达到最终的目标产物。
细胞神奇之处在于通过精心设计的代谢途径和酶催化机制,将一系列热力学上不利的化学反应与有利的反应结合起来,确保生命过程的化学平衡和能量转化,这是所有生命体共有的基础生理特征。🏳️🌈